

Buforowanie pH
Przeciwdziałanie zmianom pH przestrzeni pozakomórkowej jest bardzo istotnym elementem homeostazy, warunkującym optymalne funkcjonowanie wielu narządów. Z tego powodu nasz ‘ustrój’ zużywa mnóstwo energii na utrzymanie właściwego pH.
Podkreślam ten fakt nie bez przyczyny, gdyż chcę zwrócić uwagę na priorytety, którymi kieruje się organizm w odpowiedzi na wszystko, co ludzie ‘wyczyniają’ ze swoim ciałem (np. stosując się do zaleceń jakiejś diety-cud*). Realizując te priorytety organizm robi To-Co-Musi, a nie to, co – według nas – powinien musieć.
*) Ostatnio natknąłem się na ‘jutuba’ pewnej paleo-pani (twierdzącej, że jest dietetykiem klinicznym), która utrzymuje, że ludzki organizm nie potrafi produkować… cholesterolu (sic!). W związku z tym zaleca codzienne śniadania z 5 jaj na boczku, 600-gramowy stek na obiad i całkowitą eliminację węglowodanów złożonych.
Pomijam już fakt, że – w swej ignorancji – zaliczyła cholesterol do witamin (i na tym błędzie oparła całą swoją filozofię żywieniową), ale poprzez swoje porady nakłania ludzi do niekorzystnego dysponowania swoim zdrowiem (miażdżyca, wieńcówka, zawały, udary, tętniaki, nowotwory, cukrzyca, kamienie żółciowe, kamienie nerkowe, osteoporoza, demencja). Czasem się zastanawiam, czy to nie powinno być karalne.
Priorytety o których mowa to mechanizmy buforowania pH. Przyjrzyjmy im się. Mechanizmów jest kilka, a pierwszy z nich – buforowanie chemiczne – omówiłem w opracowaniu dotyczącym buforów osocza. W tym miejscu przypomnijmy konkluzję tamtego artykułu:
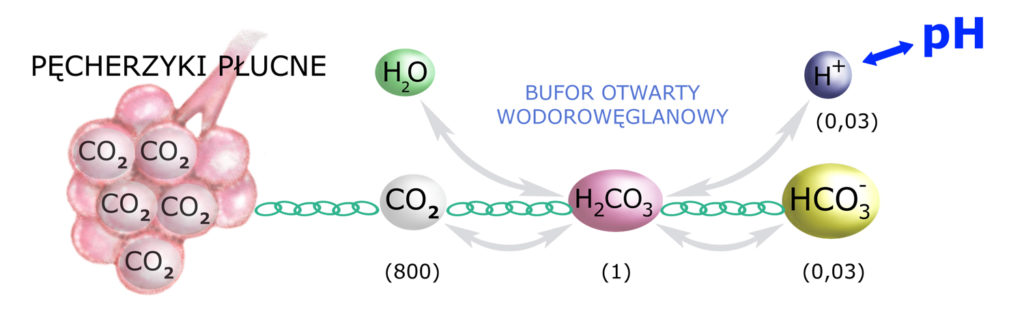
Pozostają do omówienia bufory biologiczne i fizjologiczne oraz docenienie roli rozcieńczania.
ROZCIEŃCZANIE
Chodzi o to, abyśmy zdali sobie sprawę z proporcji przestrzeni naszego organizmu. Jeśli – z dowolnych przyczyn – w naszym płynie pozakomórkowym pojawi się nadmiar kwasu lub zasady, to jest ona buforowana chemicznie (bufor wodorowęglanowy) nie tylko w osoczu krwi. Kwas lub zasada rozprowadzana jest (rozcieńczana) w całej przestrzeni pozakomórkowej – a więc również w płynie śródmiąższowym.
Pomiędzy osoczem a płynem śródmiąższowym następuje stała i dość szybka wymiana, przy czym objętość płynu śródmiąższowego jest ~3 razy większa niż objętość osocza. W związku z tym jony H+ i HCO3– dość szybko rozprzestrzeniają się w obydwu tych przestrzeniach (ich stężenia wyrównują się).
BUFOR BIOLOGICZNY
Buforem biologicznym jest przestrzeń wewnątrzkomórkowa. Wewnątrz komórek znajdują się bufory białczanowy i fosforanowy (oraz – w mniejszej ilości – wodorowęglanowy). Pojemność buforową płynu wewnątrzkomórkowego szacuje się na co najmniej tak wielką, jak cała pojemność pozakomórkowa.
Nadmiar jonów H+ z płynu pozakomórkowego może przenikać przez błony komórkowe do wnętrza komórek, przy czym – zgodnie z prawem elektroobojętności – towarzyszył mu będzie jakiś anion (np. Cl–) lub też H+ wymieni się na inny kation (np. K+). Wewnątrz komórki H+ połączy się z anionami białczanowymi i estrami organicznymi kwasu fosforowego. Przez to siła jonów H+ w płynie pozakomórkowym spadnie.
Przyjmuje się ogólnie, że ~55% jonów H+ może być buforowane przez płyn wewnątrzkomórkowy, dalsze ~30% przez płyn śródmiąższowy, a pozostałe 15-20% przez osocze krwi. To oznacza, że głównym obszarem buforowym naszego organizmu jest właśnie bufor biologiczny, czyli wnętrza naszych komórek.
BUFOR FIZJOLOGICZNY
/NERKA/
Buforowanie chemiczne zachodzi dość szybko, w ciągu kilku minut. Jeśli nie jest ono wystarczające, do gry włącza się buforowanie biologiczne, przy czym wymiana jonów pomiędzy płynem pozakomókowym a wnętrzami komórek trwa już dłużej: kilka godzin.
W opracowaniu dotyczącym równowagi wodno-mineralnej mówiliśmy o rozmieszczeniu jonów w przestrzeniach organizmu – i o tym, że głównym kationem wnętrza komórek jest potas K+. Jeśli więc buforowanie biologiczne polega na wymianie H+ i K+ (poprzez błony komórkowe), to jego efektem będzie pojawienie się większych ilości K+ w płynie pozakomórkowym. I to jest właśnie sygnał, który uruchamia buforowanie fizjologiczne.
A więc – uruchomienie buforu biologicznego (wnętrza komórek) – od razu uruchamia też bufor fizjologiczny – NERKĘ. Uzupełnijmy więc schemat buforowania o kolejny element:
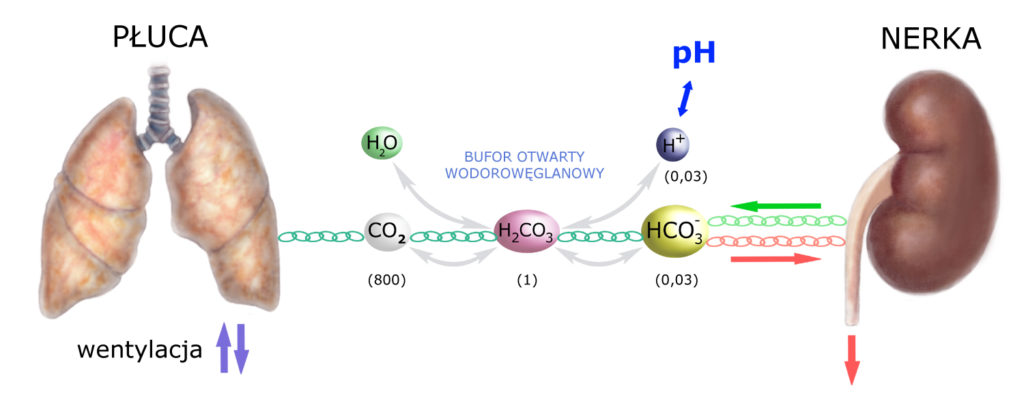
W zasadzie buforowania fizjologiczne obejmuje też układ oddechowy, a konkretnie szybkość wentylacji płuc. Nie będę się nad tym zatrzymywał, więc tylko wzmianka: płuca reagują dość szybko na zmiany stężenia CO2 we krwi. Wzrost stężenia CO2 prowadzi do przyspieszenia częstości (lub głębokości) oddechów, a więc zwiększa wentylację płuc, natomiast spadek stężenia CO2 – wentylację zmniejsza.
Regulacja ze strony układu oddechowego ma jednak swoją ograniczoną wydajność. Jeśli okazuje się ona niewystarczająca – do gry i musi wkroczyć nerka. Podczas gdy regulacja ze strony układu oddechowego ogranicza się do stężenia CO2 – nerka odpowiada za regulację stężenia wodorowęglanów (HCO3–).
Można powiedzieć, że nerka ustawicznie CZUWA nad prawidłowym stężeniem HCO3–. A więc – generalnie:
- w warunkach prawidłowych nerka wchłania HCO3– w kanalikach nerkowych i zwraca je z przesączu nerkowego do osocza krwi.
- jeśli pojawi się jakikolwiek nadmiar HCO3–, to nerka przestaje je wchłaniać i zwracać, tylko je wydala (pozwala im przedostawać się do moczu).
- jeśli jonów HCO3– jest zbyt mało (gdyż – gdzieś w organizmie – zostały zużyte do zobojętniania jakichś kwasów), to nerka odtwarza je, a więc produkuje, przywracając właściwe stężenie HCO3– w osoczu. (Podczas produkcji każdej cząsteczki HCO3– powstaje, rzecz jasna, jeden jon H+, ale ten jest od razu wydalany z moczem.)
W jaki sposób nerka radzi sobie z wydalaniem H+? Kanaliki nerkowe produkują AMONIAK (NH3), który jest akceptorem jonów H+. Drugim ważnym akceptorem H+ (choć ilościowo mniejszym) są jony fosforanowe (HPO42-). Akceptorami są też (w ilościach znikomych) kreatynina i kwas moczowy.

Ważne jest, aby zrozumieć jak to działa.
Wyobraźmy sobie, że do naszego organizmu dostał się jakiś dodatkowy, niepotrzebny nam kwas. Generalnie – kwas ten będzie się zawsze składał z jonu H+ i jakiejś zasady anionowej, którą symbolicznie zapiszmy jako A–. Do układu tego natychmiast dobiorą się nasze jony wodorowęglanowe:
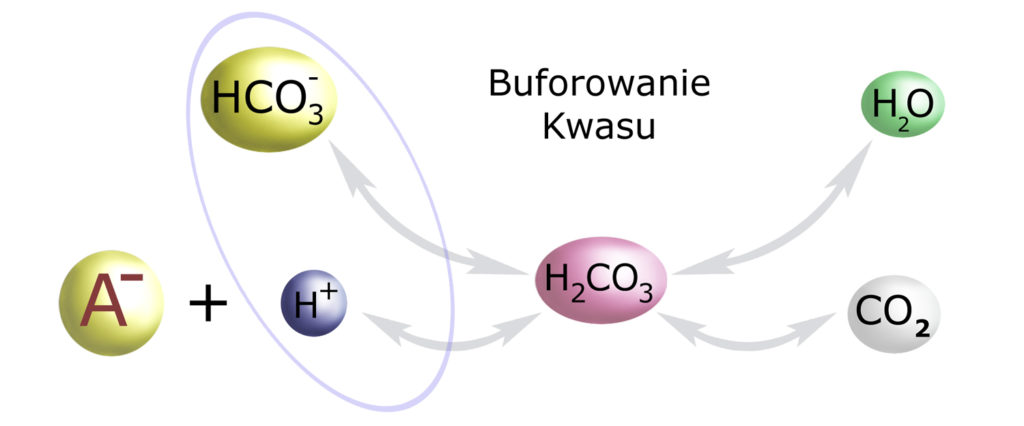
Jak widać, wodorowęglan łączy się z wodorem, neutralizując go. Oba znikają, gdyż zamieniają się z kwas węglowy, a ten dalej w wodę i dwutlenek węgla (którego pozbędą się nasze płuca). Pozostaje jedynie zasada anionowa, którą ostatecznie wydali nerka – kiedy tylko ta do niej trafi.
Jako że – buforując kwas – znikł nam wodorowęglan, więc nerka musi go odtworzyć. Produkując wodorowęglan – powstaje jon H+, który nerka wydala – łącząc go z amoniakiem lub fosforanem. Dorysujmy to do obrazka:
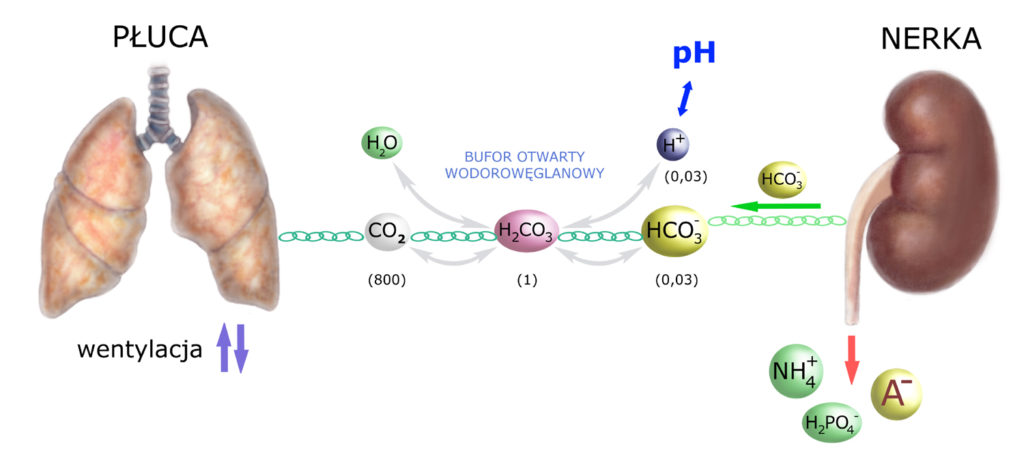
Powyższy obrazek dotyczy buforowania kwasu. Jeśli nerka buforuje zasadę, to wydala HCO3–, przy czym – aby została zachowana zasada elektroobojętności moczu – wraz z wodorowęglanem nerka wydala też kation Na+. Taka sytuacja doprowadza do zwiększenia ilości wydalanego moczu (gdyż za sodem – tonicznie – podąża woda), co jest wskazówką zwiększonego wydalania elektrolitów.
Tak więc – podsumowując – rola nerki sprowadza się do alkalizacji lub zakwaszania moczu. Zakwaszając mocz, nerka jednocześnie odkwasza organizm: produkuje HCO3– i zwraca je do osocza.
* *
A teraz pytanie za tysiąc punktów:
Skąd Wziął Się ten Kwas w Naszym Organizmie?
Chodzimy sobie, bogu ducha winni, po świecie, oddychamy, gramy w bierki, rozważamy o Arystotelesie lub ‘dupie Maryni’… aż tu nagle KWAS w organizmie. Skąd się wziął? Przecież nasz organizm nie mógł, nagle i niespodziewanie – sam z siebie – go wyprodukować. Skąd by go wziął?
To musi być coś z zewnątrz, coś z otoczenia. A jakie są wrota kontaktu naszego organizmu z otoczeniem? Kwas wniknął przez skórę? Nawdychaliśmy się go przez płuca? Czy może, po prostu…
Zjedliśmy Go!
Organizm ważący 60, 80 czy 100 kg przepuszcza przez siebie w ciągu roku ~1,5 tony (a czasem i więcej) związków chemicznych zwanych jedzeniem i piciem. Pomijając czystą wodę, wszystko co jemy lub pijemy ma jakiś odczyn – kwasowy lub zasadowy.
Owszem, zakwaszenie organizmu może mieć różne przyczyny (np. wysiłek fizyczny produkujący kwas mlekowy w naszych mięśniach) ale – generalnie – za zakwaszenie odpowiada to, co najczęściej codziennie wkładamy do ust. Czyli nasza dieta, rozumiana jako permanentny sposób żywienia.
Najczęstszym źródłem zakwaszenia organizmu, z którym muszą potem radzić sobie nerki, są mocne kwasy – siarkowy i fosforowy – metabolity przemiany białkowej. (Dlatego przy chorobach nerek ‘dietą z wyboru’ jest dieta niskobiałkowa). W warunkach normalnych powstają też w organizmie mocne kwasy organiczne (mlekowy, cytrynowy, pirogronowy, acetooctowy i β-hydroksymasłowy), ale jest ich niewiele a ich przemiana szybka, więc praktycznie nie mają wpływu na równowagę kwasowo-zasadową.
A więc… BIAŁKO.
A konkretnie – jego nadmiar.
I tu – po raz kolejny – trzeba się zmierzyć z iście religijnym dogmatem dietetycznym, który osobiście nazywam supergumnoprawdą: ‘im więcej białka tym lepiej’. Bo przecież białko jest dla nas… dobre. Cóż…
Ciepło też jest dobre – wykąp się we wrzątku.
Ogólna zasada jest taka:
Przy diecie białkowej – kwaśność moczu rośnie (nerka odkwasza organizm).
Przy diecie jarskiej – kwaśność moczu maleje.
A teraz pora na przykład.
We wstępie wspomniałem o paleo-uroczej, spontanicznie uśmiechniętej pani, która zaleca dietę pozbawioną węglowodanów złożonych. Jako że w tym opracowaniu rozważamy przypadki kwasicy, przyjrzyjmy się bliżej temu, co dzieję się w jej organizmie – właśnie pod kątem zakwaszenia. Przy okazji – rozpatrzę też przypadek cukrzycy, gdyż jest więcej niż pewne, że już niedługo nasza paleo-urocza na nią zachoruje. Tak (!), gdyż kłopoty z glikemią nie są przyczyną cukrzycy, ale jej skutkiem. Przyczyną są zmiany receptora insulinowego, który staje się mało wrażliwy na normalne stężenia insuliny (insulinooporność) pod wpływem diet obfitujących w tłuszcze nasycone = zwierzęce (można wręcz powiedzieć: ‘tłuszcze paleo’).
Kwasica Ketonowa
(Cukrzycowa)
W przypadkach głodzenia, cukrzycy czy stosowania diet proteinowych dochodzi do tzw. kwasicy ketonowej. Nie dostarczając organizmowi węglowodanów (które rozkładają się do glukozy – naszego głównego substratu energetycznego), zmuszamy organizm do produkowania energii z innego źródła: z naszej tarczy antykryzysowej, czyli z tkanki tłuszczowej.
Gospodarkę energetyczną organizmu kontrolują dwa hormony produkowane przez trzustkę: INSULINA i GLUKAGON, przy czym insulina jest hormonem anabolicznym (zwiększa zapasy), a glukagon – katabolicznym (trwoni zapasy).

Osłabiona (i z biegiem czasu coraz słabsza) wrażliwość receptora insulinowego na insulinę powoduje, że glukoza ma kłopoty z przedostawaniem się do komórek, aby zostać zmagazynowana pod postacią glikogenu (w wątrobie i mięśniach) lub tłuszczu (w tkance tłuszczowej). Pozostaje więc we krwi w dużych stężeniach (hiperglikemia).
Nieskuteczność insuliny sprawia, że przewagę zyskuje glukagon, który pogłębia tragedię. Powodując rozpad glikogenu do glukozy i syntezę glukozy z aminokwasów (których jest mnóstwo, bo glukagon przyspiesza rozpad białek) – pogłębia już istniejącą hiperglikemię.
Pozostająca poza komórkami glukoza sprawia, że nie mogą jej one wykorzystywać jako materiału energetycznego. Glukagon jednak powoduje również rozpad tłuszczów, więc we krwi pojawia się mnóstwo wolnych kwasów tłuszczowych (WKT), które mogą stanowić ‘paliwo zastępcze’.
Jednak (!) na skutek zwiększonej podaży WKT do wątroby – dochodzi do szybkiego wyczerpania się pewnych metabolitów umożliwiających pełne spalanie WKT (do CO2 i H2O). Nagromadzony Acetylo-CoA musi więc spalać się w sposób niepełny – do tzw. związków ketonowych. Są nimi kwas acetooctowy i kwas β-hydroksymasłowy.
Zdolność produkowania związków ketonowych z WKT posiada w zasadzie tylko wątroba. Sama z nich jednak nie korzysta, tylko uwalnia je do krwi, gdzie stają się dobrym materiałem energetycznym dla nerek, mięśni szkieletowych i mięśnia sercowego.
Opisana powyżej sytuacja jest charakterystyczna dla cukrzycy, lecz zwróć uwagę na to, że – pomijając hiperglikemię – ten schemat dotyczy też zarówno głodówki jak i diety proteinowej, gdyż w obu tych przypadkach komórki organizmu pozbawione są dostaw glukozy. (Zapasy glukozy zmagazynowanej pod postacią glikogenu wyczerpują się w ciągu doby.) Tutaj również zaczyna królować glukagon uwalniający rezerwy energetyczne: inicjuje rozpad lipidów do WKT (z których powstają związki ketonowe) i rozpad białek do aminokwasów (z których powstaje glukoza – niezbędna dla krwinek czerwonych i prawidłowego funkcjonowania ośrodkowego układu nerwowego).
We wszystkich tych przypadkach mamy do czynienia z sytuacją nienaturalną – czerpiemy energię z ‘paliwa zastępczego’ którym są KWASY (acetooctowy i β-hydroksymasłowy). Stan taki nie jest dla nas niebezpieczny, jeśli jest krótkotrwały. Kiedy jednak przedłuża się w nieskończoność, stężenie kwasów we krwi rośnie wielokrotnie, zaczynamy je wydalać z moczem a nasz oddech ‘zionie’ acetonem – co oznacza, że już buforujemy.
W dietach proteinowych do zakwaszenia pochodzącego od związków ketonowych należy dodać zakwaszenie pochodzące od białek mięsa, nabiału i jaj – zżeranych przez ‘smuklącą się’ paleo-celebrytkę lub ‘robiącego rzeźbę’ paleo-osiłka.
Przy okazji (muszę, bo to ważne):
Co się teraz dzieje z wapniem?
Opisując bufory krwi wspomniałem, że prócz dwóch głównych (wodorowęglanowego i hemoglobinowego) w osoczu są jeszcze bufory białczanowe, fosforanowe i kwasów organicznych (dające łącznie ~12% pojemności buforowania). Wapń występuje w osoczu w dwóch postaciach: wolnej (zjonizowanej) i jako Ca2+ związany – m. in. właśnie ze wspomnianymi buforami (z anionami cytrynianów, fosforanów i białek, głównie frakcji albumin).
W chwili zakwaszenia organizmu związki te odłączają Ca2+ i przyłączają H+ – bo buforują krew (zmniejszając ilość wolnego H+ w osoczu)! Oznacza to, że nagle we krwi jest zbyt dużo wolnego Ca2+ – więc nerki zaczynają go wydalać w dużych ilościach (co często prowadzi do powstawania kamieni nerkowych).
Gdy organizm chwilowo upora się z zakwaszeniem bufory znów zwiążą Ca2+, a więc teraz będzie go za mało w formie wolnej. Ubytki utraconego wapnia uzupełniane są… z kości – pogłębiając osteoporozę! Nie z posiłków, bo wszystkie posiłki są paleo-zakwaszające, co (patrz wyżej) powoduje, że podczas trawienia poziom wolnego Ca2+ we krwi jest wysoki, a więc jelito go nie wchłania.)
Sam fakt, że zaczynamy buforować naszą cud-dietę – świadczy o tym, że nasz organizm uznaje ten stan za niepożądany. Organizm chce pozbyć się kwasów i przywrócić homeostazę – a to znaczy, że w niej NIE JEST. Diety proteinowe, diety ‘keto’ – są po prostu sztucznym wyprowadzaniem organizmu z homeostazy, a więc ze stanu zdrowia. A skoro ‘ze zdrowia’, to – w co?
Wracając do cukrzycy:
Zauważ, że wstrzykiwanie sobie sztucznie produkowanej insuliny w dużych dawkach wcale nie jest leczeniem cukrzycy, tylko zaleczaniem jej objawów (bombardujemy słabe receptory insulinowe nadmiarem insuliny, aby wreszcie zareagowały). Leczeniem cukrzycy jest przywracanie sprawności receptorom. I to się da zrobić.
Cukrzyca jest chorobą dietozależną (!), a takie choroby można wyleczyć jedynie dietą. Istnieją lekarze, którzy dokładnie to właśnie robią. Należy do nich np. dr John McDougall, o którym już pisałem i zapewne jeszcze wielokrotnie będę pisał.
McDougall twierdzi, że cukrzyca jest chorobą w pełni uleczalną, a niektórzy jego pacjenci odstawiają insulinę już po 2-3 tygodniach. Cóż takiego McDougall daje swoim pacjentom do jedzenia? Odstawia produkty zwierzęce (w 100%), zamieniając je na roślinne – węglowodany złożone, takie jak ziemniaki lub ryż (przez cały czas monitorując zdrowie pacjenta i interweniując kiedy trzeba).
Jeśli masz kłopoty z insulinoopornością lub jesteś cukrzykiem ‘pełną gębą’ – warto poprosić twojego lekarza, aby się tym zainteresował. A nóż trafisz na doktora z otwartym umysłem i bez kompleksów. To się zdarza.