

HOMEOSTAZA – PRAWO ELEKTROOBOJĘTNOŚCI
We wprowadzeniu do artykułów o homeostazie wspomniałem o wzajemnych zależnościach pomiędzy wieloma parametrami fizykochemicznymi naszego organizmu. Już sama zmiana temperatury ciała – choćby pod wpływem gorączki lub nadmiernych upałów – pociąga za sobą łańcuch regulacji.
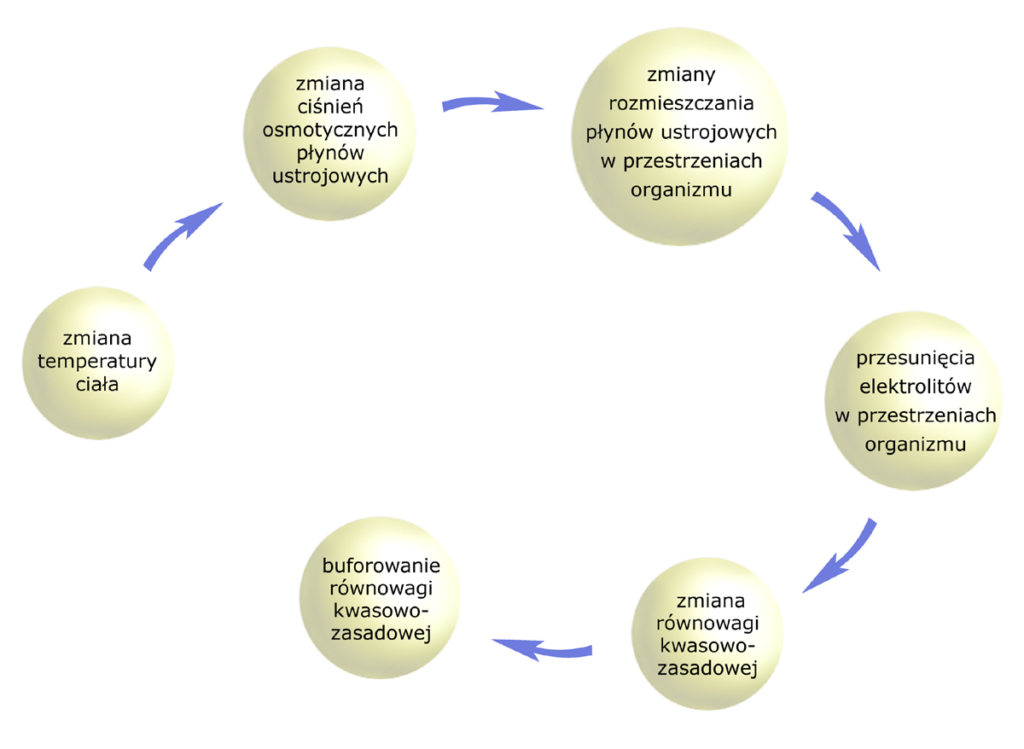
W dzisiejszym opracowaniu przyjrzymy się równowadze ładunków elektrycznych, a więc równowadze wodno-elektrolitowej (wodno-mineralnej). Podlega ona jednemu z trzech praw zawiadujących homeostazą organizmu. Jest nim…
Prawo ELEKTROOBOJĘTNOŚCI (+/-)
Suma ładunków dodatnich jest równa sumie ładunków ujemnych
A więc: suma stężeń kationów (jonów o ładunku dodatnim) jest równa sumie stężeń anionów (jonów o ładunku ujemnym) w każdej przestrzeni naszego organizmu.
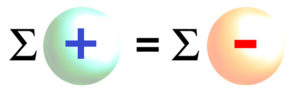
Duża grecka litera sigma symbolizuje sumę.
Chcąc pojąć czym jest i jak działa równowaga wodno-elektrolitowa, najpierw musimy się przyjrzeć tzw. przestrzeniom organizmu. Przedstawmy je za pomocą schematu:
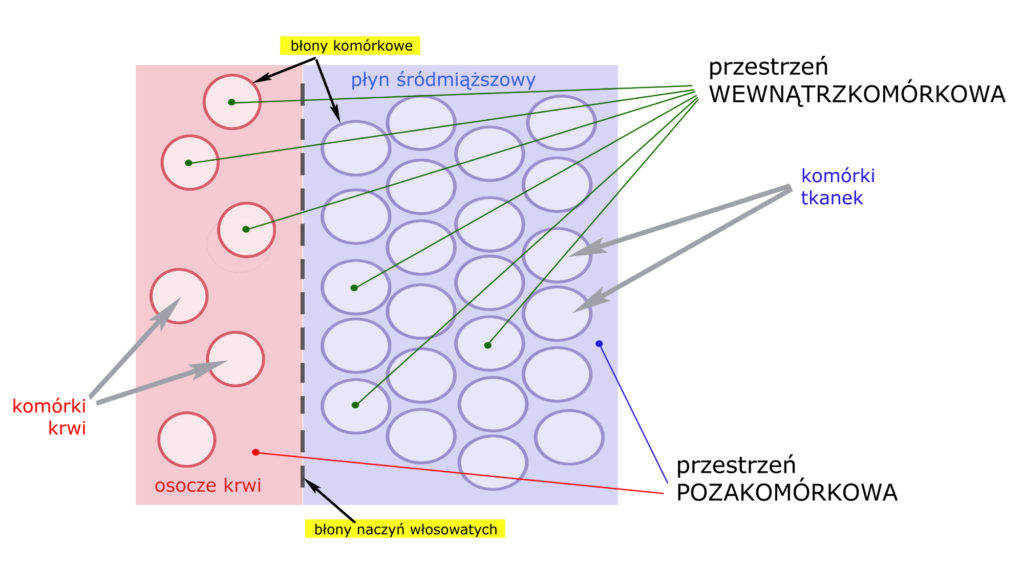
Jak widać, generalny podział jest prosty. Dwie główne przestrzenie to wewnątrzkomórkowa i pozakomórkowa. Przestrzeń wewnątrzkomórkowa zawiera 2/3 płynów organizmu. Przestrzeń pozakomórkową (1/3 płynów organizmu) stanowią – przede wszystkim – osocze krwi i płyn śródmiąższowy tkanek, oddzielone od siebie jedynie błonami* naczyń włosowatych.
*) Szczególna uwagę zwróćmy właśnie na błony biologiczne – w szczególności błony naczyń włosowatych i błony komórkowe.)
Błony naczyń włosowatych są bardzo słabą barierą, a więc znakomita większość substancji drobnocząsteczkowych rozpuszczonych w osoczu krwi – z łatwością przenika do płynu śródmiąższowego i vice versa. Następuje tu dość szybka i wydajna wymiana, można więc powiedzieć, że stężenia substancji drobnocząsteczkowych w osoczu i w całym płynie śródmiąższowym są w zasadzie takie same.
Zupełnie inna sytuacja ma miejsce na granicy przestrzeni wewnątrz- i pozakomórkowej. Barierę dla tych przestrzeni stanowią błony komórkowe, będące bardzo silną przeszkodą dla wszystkiego, co nie jest wodą.
Każda komórka naszego organizmu jest częścią jakiejś tkanki, a więc posiada specyficzne zadanie do wykonania. Wymaga to utrzymywania określonego środowiska wewnątrz komórki. Właśnie dlatego błony komórkowe wyposażone są w narzędzia umożliwiające precyzyjną kontrolę tego, co przez nie przenika.
Oczywiście przedstawiony powyżej schemat jest mocno uproszczony. Pominąłem w nim tę część przestrzeni pozakomórkowej, której zawartość jest słabo wymienialna z osoczem. Dotyczy to np. płynów tkanki łącznej, chrząstek i kości, oraz całej tzw. przestrzeni transcelularnej *. Niemniej ograniczę się do modelu uproszczonego, gdyż jest wystarczający dla zrozumienia zasad równowagi wodno-elektrolitowej
*) Przestrzeń transcelularna jest szczególnym rodzajem przestrzeni. Charakteryzuje się tym, że komunikuje się ona bezpośrednio – z wnętrzami komórek.
Przestrzeń transcelularną (transkomórkową) stanowią płyny światła przewodu pokarmowego, dróg żółciowych i trzustkowych, płyn dróg moczowych i płyny jam ciała (np. płyn mózgowo-rdzeniowy, osierdziowy, otrzewnowy, opłucnej, komór oka).
Zanim zajmę się elektrolitami i ich rozmieszczeniem w przestrzeniach organizmu, wypada jeszcze wyjaśnić jedną zagadkę: Jak to się dzieje, że przestrzeń pozakomórkowa – mimo iż jest tak jednorodna pod względem zawartości substancji w niej występujących – jakimś cudem daje sobie radę w wymianie metabolitów.
Rozwiązaniem tego problemu jest tzw. równowaga Starlinga. Najlepiej ją zrozumieć analizując rysunek znajdujący się nieco poniżej. Przedstawia on schemat budowy naczyń włosowatych oraz, na powiększeniu, układ: osocze / błona naczynia włosowatego / płyn śródmiąższowy.
Tętnice dzielą się na mniejsze tętnice, te na tętniczki, te na jeszcze mniejsze tętniczki i tak dalej, aż w końcu – na poziomie tkanki – powstają super cienkie kapilary, zwane naczyniami włosowatymi. Tu następuje wymiana substancji drobnocząsteczkowych pomiędzy osoczem i płynem śródmiąższowym. Po wymianie naczynie włosowate zaczyna łączyć się z innymi naczyniami w maluteńkie żyłeczeńki, te w coraz to większe żyłeczki i w końcu w żyły.
Zadaniem krwi tętniczej jest dostarczanie do tkanek wszystkiego, czego one potrzebują, krew żylna natomiast odprowadza z tkanek niepotrzebne im już produkty przemiany materii. Cała wymiana (co jest istotą sprawy) zachodzi więc na poziomie błon naczyń włosowatych, dlatego na nich się skupmy:
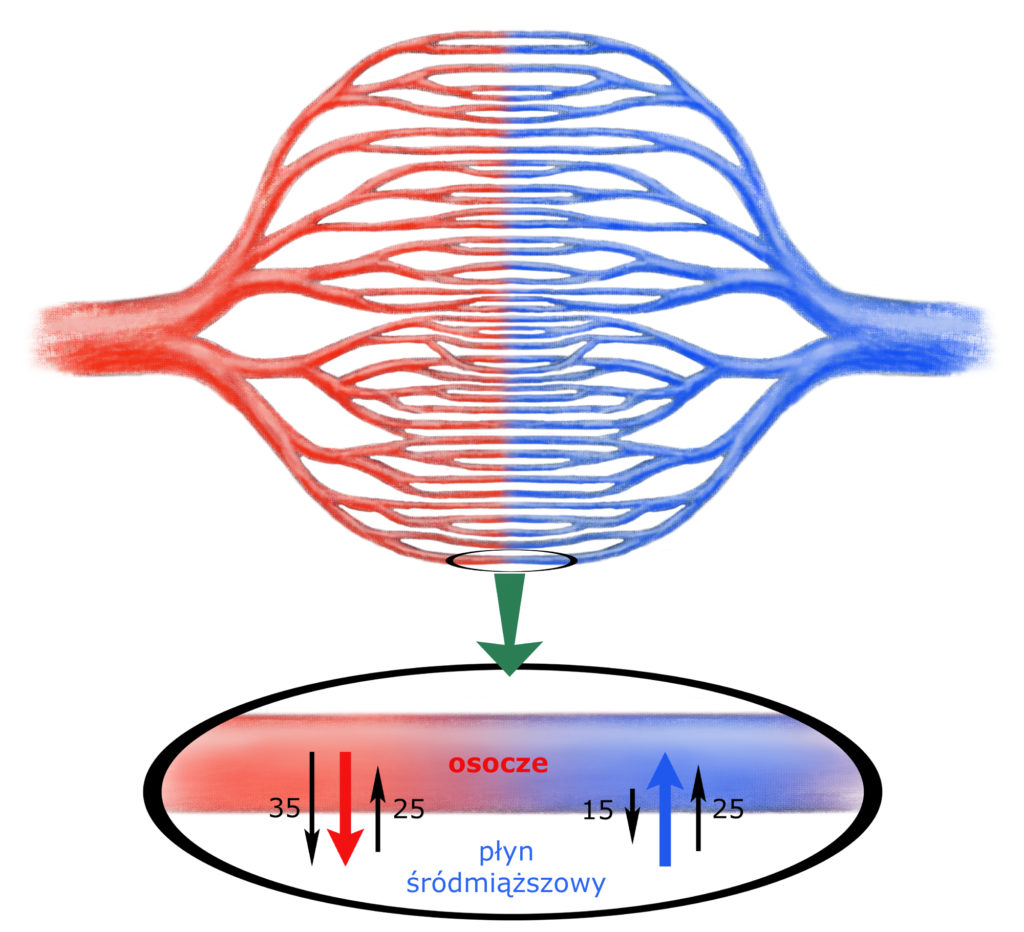
Najważniejszą sprawą jest uzmysłowienie sobie, że na poziomie tkankowym, a więc na układ: osocze <=> płyn śródmiąższowy – działają dwa rodzaje ciśnień:
- hydrostatyczne – wypychające wodę z osocza do płynu śródmiąższowego
- osmotyczne (toniczne) – wsysające wodę z płynu śródmiąższowego do osocza
Zacznijmy od ciśnienia tonicznego.
Skąd bierze się ciśnienie osmotyczne w układzie osocze <=> płyn śródmiąższowy? Otóż osocze zawiera dużo wielkocząsteczkowych białek, natomiast płyn śródmiąższowy – mało. Błona naczynia włosowatego jest łatwo przepuszczalna dla wody i substancji drobnocząsteczkowych. Wielkocząsteczkowe białka przez cały czas pozostają więc wewnątrz naczynia włosowatego – budując różnicę ciśnień osmotycznych pomiędzy osoczem krwi a płynem śródmiąższowym tkanek. Ciśnienie to wynosi 25 mm Hg i jest stałe na całej długości naczynia włosowatego.
Teraz ciśnienie hydrostatyczne:
W miarę podziału tętnic panujące w nich ciśnienie hydrostatyczne się zmniejsza – podobnie jak w wężu ogrodowym, który rozdzielamy na odnogi, aby zaopatrzyć w wodę wiele grządek: każde kolejne odgałęzienie przejmuje tylko część ciśnienia głównego zaworu wody. Każde naczynie włosowate posiada więc tylko maleńką cząstkę ciśnienia hydrostatycznego, w które nasze serce zaopatruje cały układ naczyniowy. Na samym początku naczynia włosowatego wynosi ono 35 mm Hg.
Spójrzmy – co się dzieje na samym początku kapilary. (Ten jej początkowy odcinek nazywamy częścią tętniczą naczynia włosowatego.) Ciśnienie wypychające wodę z naczynia (hydrostatyczne) wynosi tu 35 mm Hg, a wsysające wodę do naczynia (osmotyczne) tylko 25 mm Hg. Mamy więc przewagę siły wypychającej wodę z naczynia – równą 10 mm Hg. Właśnie dlatego w początkowym odcinku naczynia włosowatego woda i rozpuszczone w niej substancje drobnocząsteczkowe (!) – przenikają z osocza do płynu śródmiąższowego.
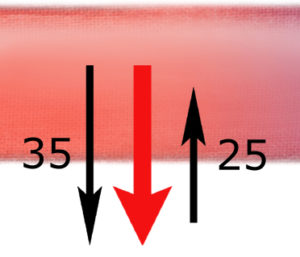
W miarę uciekania wody z naczynia – spada panujące w nim ciśnienie hydrostatyczne do 15 mm Hg, natomiast ciśnienie osmotyczne pozostaje niezmienione (25 mm Hg).
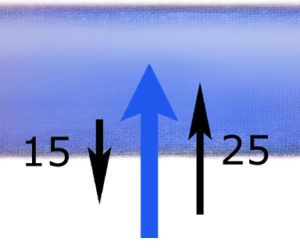
W drugiej części kapilary (części żylnej naczynia włosowatego) mamy więc sytuację odwrotną: tutaj przeważa siła wsysająca wodę do naczynia – również o 10 mm Hg (osmotyczne – wciąż 25, ale hydrostatyczne – już tylko 15). W efekcie w drugiej części naczynia włosowatego – woda i rozpuszczone w niej substancje drobnocząsteczkowe (!) przenikają z płynu śródmiąższowego do osocza.
Łącząc wszystko w całość:
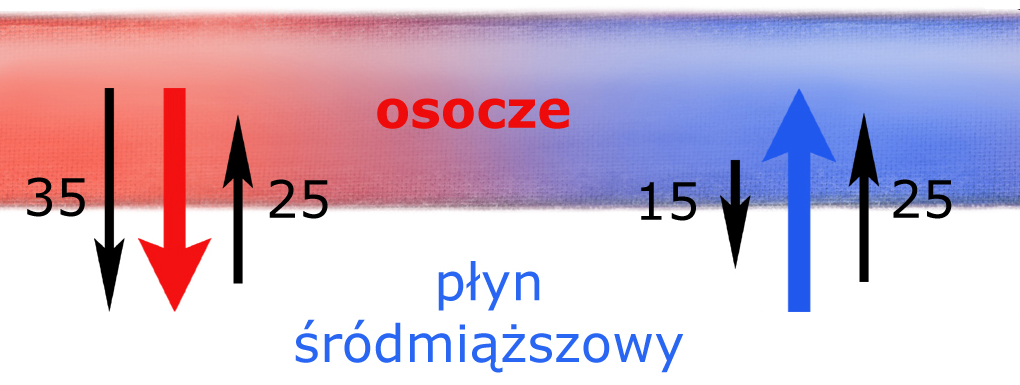
A) W części tętniczej naczynia włosowatego woda z krwi tętniczej, wraz z potrzebnymi tkankom rozpuszczonymi w niej substancjami drobnocząsteczkowymi (tlen, sód, potas, chlorki, glukoza, wodorowęglany, itd.) – przemieszcza się do płynu śródmiąższowego tkanek.
B) W części żylnej naczynia włosowatego prawie identyczna* objętość wody wraca do naczynia, z tym że tym razem jest to woda z płynu śródmiąższowego, wraz z jej substancjami drobnocząsteczkowymi (m.in. z mniejszym stężeniem glukozy a większą zawartością dwutlenku węgla i innych produktów przemiany z komórek.)
*) Napisałem ‘prawie identyczna’ gdyż jest jej nieco mniej. Jednak ta część wody osocza, która pozostała w płynie śródmiąższowym – i tak ostatecznie wraca do naczyń krwionośnych pod postacią chłonki (transportowana układem chłonnym do żył podobojczykowych).
Tak właśnie działa tzw. równowaga Starlinga na poziomie błon naczyń włosowatych. Dzięki niej zawartość osocza nieustannie wymienia się z zawartością płynu śródmiąższowego.
* *
Rzecz jasna – jak można się domyślać – tzw. mikrokrążenie (a więc filtracja i reabsorpcja płynu przez ściany naczyń włosowatych) jest o wiele bardziej skomplikowanym procesem.
Części tętnicze naczyń włosowatych uzbrojone są w tzw. zwieracze przedwłośniczkowe zbudowane z mięśni gładkich zdolnych do zamykania i otwierania światła naczyń. Mają one zdolność autoregulacji przepływu krwi; pulsując, zamykają i otwierają poszczególne naczynia włosowate tak, że zwykle otwarte jest ~25% naczyń. W naczyniach otwartych przeważa ciśnienie hydrostatyczne, a więc filtracja z osocza do płynu śródmiąższowego, natomiast w zamkniętych przeważa ciśnienie osmotyczne, czyli reabsorpcja z płynu śródmiąższowego do osocza.
Zwieracze przedwłośniczkowe są ponadto bardzo wrażliwe na czynniki hormonalne, dzięki czemu gruczoły dokrewne mogą wpływać na efekt wymiany płynów, regulując rozmieszczenie wody w przestrzeniach organizmu, a więc m. in. wypełnienie łożyska naczyniowego (ciśnienie krwi).
Regulacja zachodzi też na poziomie ścian naczyń włosowatych (budowa ciągła, nieciągła i okienkowata). W ich śródbłonku występują nitki białek kurczliwych (aktyna i miozyna), których skurcz wywoływany jest napływem jonów Ca2+. W wywołaniu reakcji tych białek pośredniczą swoiste receptory śródbłonka, wrażliwe na niektóre kininy osocza. (Reagują też na substancje uwalniane z uszkodzonych tkanek; powoduje to wzrost przepuszczalności naczyń włosowatych, prowadząc do obrzęku zapalnego).
Wypada też wspomnieć o tzw. zespoleniach tętniczo-żylnych, dzięki którym krew może omijać mikrokrążenie, wpływając na termoregulację (szczególnie niektórych obszarów skóry).
* *
Rozmieszczenie Elektrolitów;
Główne Kationy i Aniony Organizmu
Przeanalizowaliśmy już co dzieje się na poziomie błon naczyń włosowatych. Pozostają błony komórkowe, dzielące organizm na przestrzenie wewnątrz- i pozakomórkową.
Wspominałem, że komórki poszczególnych tkanek, pełniąc odmienne funkcje w organizmie, indywidualnie kontrolują przepływ substancji przez swoje błony. Istnieje jednakże (!) pewien generalny schemat rozmieszczenia substancji drobnocząsteczkowych w przestrzeniach wewnątrz- i pozakomórkowej. Oto on:
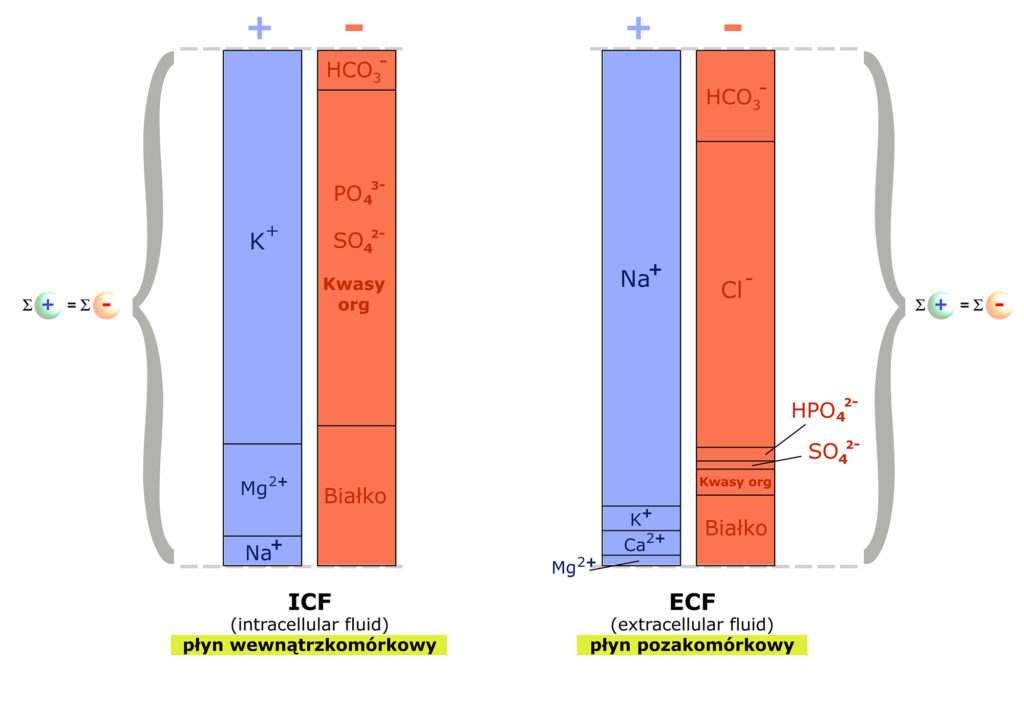
Zwróć uwagę na identyczną wysokość słupków (oznaczona klamrami): w obu przestrzeniach zachowane jest prawo elektroobojętności – sumy kationów i anionów są sobie równe.
Już pierwszy rzut oka na powyższy schemat pokazuje zasadnicze różnice w składzie jonowym wnętrza komórek i przestrzeni pozakomórkowej – generalną prawdę dotyczącą rozmieszczenia elektrolitów w naszym organizmie.
Głównym kationem wnętrz komórek jest potas (K+), a aniony pochodzą od substancji organicznych. Jest to – przede wszystkim – jon fosforanowy (PO4-3) oraz aminokwasy.
Głównym kationem przestrzeni pozakomórkowej, a więc osocza i płynu śródmiąższowego tkanek – jest sód (Na+), a głównym pozakomórkowym anionem – chlor (Cl–).
Stan taki – jak się domyślasz – nie jest przypadkiem. Nasz organizm bardzo pieczołowicie dba o to, aby rozmieszczenie elektrolitów wyglądało właśnie tak, jak przedstawiono to powyżej. Taki ‘układ’ jonów jest stanem idealnym i docelowym. Jest częścią HOMEOSTAZY – podstawą stałości środowiska fizyko-chemicznego panującego wewnątrz naszego organizmu. Wszelkie odstępstwa od tego stanu skutkować będą całymi łańcuchami różnorakich reakcji ze strony naszego organizmu, mających tylko jeden cel: przywrócenie idealnego stanu docelowego.
Przyszła pora na najtrudniejsze z pytań – pytania elementarne (z niewiadomych powodów zadawane przez nas głównie w wieku lat czterech): PO CO? i DLACZEGO? Najzabawniejsze jest to, że można tygodniami czytać książki i podręczniki medyczne (takie jak ‘Fizjologia Człowieka’ Traczyka i Trzebskiego albo ’Biochemia’ Harpera) i nie znaleźć prostej i jednoznacznej odpowiedzi na to pytanie.
No więc po co nam ten stan? Dlaczego w ogóle istnieje?
Otóż takie proporcje elektrolitów – poza tym, że stanowią idealne środowisko dla reakcji biochemicznych – są wskaźnikiem właściwego rozmieszczenia wody w poszczególnych przestrzeniach ciała, w szczególności wskaźnikiem objętością krwi. Dowolne zachwianie stałości środowiska wewnętrznego (homeostazy) – prowadzi w pierwszej kolejności do zmian w rozmieszczeniu wody, o czym nasz organizm dowiaduje się szybko, monitorując stężenia swoich elektrolitów – przede wszystkim sodu i potasu.
Woda wiązana jest przecież osmotycznie:
– przez sód – w przestrzeni pozakomórkowej
– przez potas – w przestrzeni wewnątrzkomórkowej
Jeśli więc woda przesuwa się pomiędzy przestrzeniami organizmu – oznacza to, że zmieniły się warunki osmotyczne, czyli stężenia sodu i potasu.
Zmiany bilansu wodnego mogą bardzo szybko doprowadzić do naszej śmierci, dlatego kontrola rozmieszczenia wody – równoznaczna z kontrolą poziomów sodu i potasu – jest ekstremalnie ważnym elementem homeostazy.
Jak to się dzieje, że elektrolity rozmieszczone są tak nierównomiernie pomiędzy przestrzeniami wewnątrz- i pozakomórkową? Przyczyną są:
Układy Transportujące Jony
poprzez błony komórek
Mamy tu do czynienia z transportem biernym, zachodzącym bez nakładu energii metabolicznej oraz z transportem czynnym (aktywnym), wykorzystującym energię i działającym przeciw gradientom stężeń.
Transport bierny
Zachodzi on zgodnie z gradientem stężeń i wykorzystuje zjawiska dyfuzji prostej oraz dyfuzji ułatwionej (zwanej też transportem nośnikowym).
1. Cząsteczki rozpuszczalne w tłuszczach przechodzą przez błonę komórkową w trybie dyfuzji prostej (od stężenia większego do mniejszego).
Należą tu m.in. cząsteczki O2 i CO2.
2. Cząsteczki nierozpuszczalne w tłuszczach wykorzystują dyfuzję ułatwioną: po jednej stronie błony komórkowej łączą się one z nośnikiem – białkiem transportującym, stanowiącym integralną część błony. W efekcie tego połączenia powstaje nietrwały kompleks cząsteczka-białko, który jest już rozpuszczalny w tłuszczach. Po drugiej stronie błony kompleks ulega dysocjacji – cząsteczka odłącza się od białka.
W ten sposób przechodzą przez błonę m.in. glukoza i aminokwasy.
3. Cząsteczki obojętne lub mające ładunek ujemny przechodzą przez błonę bez trudu poprzez kanały jonowe (białka spełniające rolę porów w błonie komórkowej).
Należą tu woda, mocznik lub aniony Cl–.
4. Cząsteczki o ładunku dodatnim napotykają jednak na małą przeszkodę. Otóż w otoczeniu kanałów jonowych znajdują się naładowane dodatnio białka oraz kationy Ca2+, które odpychają cząsteczki o ładunku dodatnim. Mimo to dyfuzja zachodzi, gdyż wnętrze komórek jest naładowane ujemnie.
Należą tu m.in. kationy Na+ i K+.
Transport czynny (aktywny)
Wydajność transportu biernego nie jest w stanie doprowadzić do takiej dysproporcji w rozmieszczeniu jonów, jaką obserwujemy pomiędzy wnętrzem komórek i przestrzenią pozakomórkową.
Wysokie stężenie Na+ poza komórką utrzymywane wbrew siłom elektrycznym (ujemnie naładowane wnętrze komórki) oraz dyfuzyjnym (mało sodu wewnątrz komórki), jak również gradient stężeń K+ wewnątrz i poza komórką – muszą być efektem czynnego WYRZUTU jonów sodowych z wnętrza komórki i aktywnego WRZUTU potasu do jej wnętrza.
Takie właśnie działanie ma tzw. pompa sodowo-potasowa, wykorzystująca energię metaboliczną organizmu. Jej działanie nadzoruje enzym błony komórkowej, zwany ATP-azą Na+–K+ (adenozynotrójfosfataza aktywowana przez jony sodu i potasu). Enzym ten jednocześnie wrzuca 2 kationy potasu do komórki i wyrzuca z niej 3 kationy sodu, czerpiąc energię z reakcji odczepienia fosforanu od ATP (w obecności magnezu):
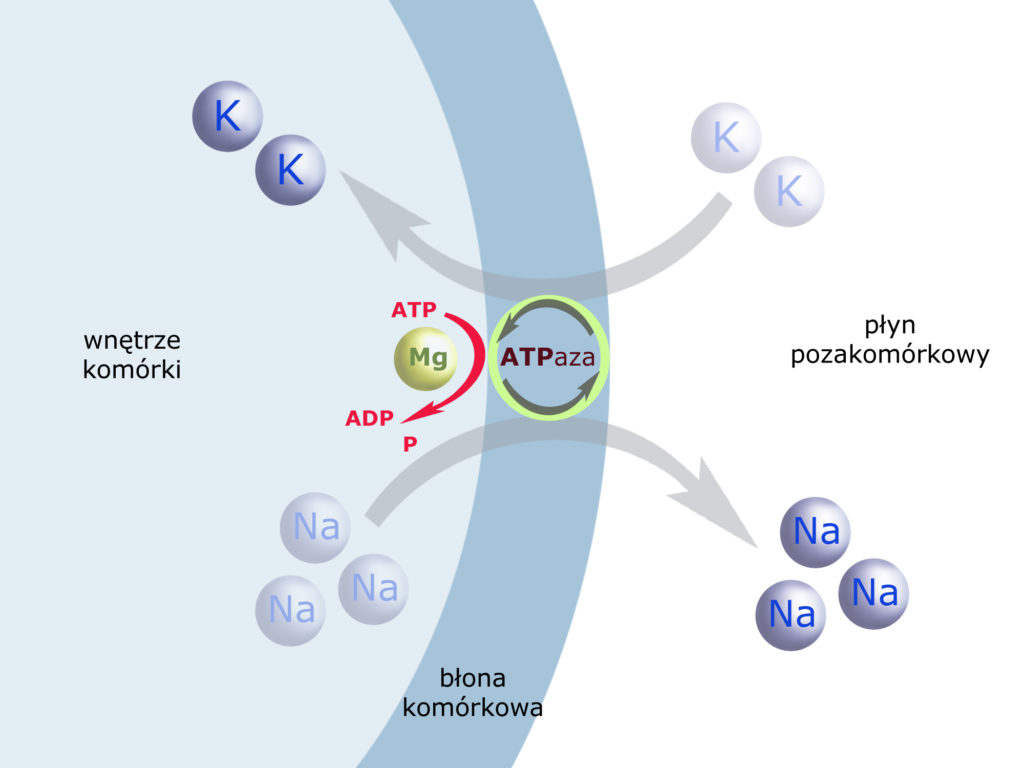
To właśnie aktywność pompy sodowo-potasowej sprawia, że wnętrze komórki zawsze posiada ujemny potencjał elektrochemiczny (wrzucając dwa kationy – jednocześnie wyrzuca trzy). Nie jest to złamanie prawa elektroobojętności, a – wręcz przeciwnie: wykorzystanie go do utworzenia i utrzymywania 'siły’ przyciągającej kationy do komórki.
Duże znaczenie ma też ATP-aza Ca2+, transportująca jony wapniowe, czy ATP-aza H+, zwana pompą protonową. W błonach działają też inne mechanizmy transportu jonów, choćby wymieniacze jonowe (sodowo-wapniowe, sodowo-wodorowe, chlorkowo-wodorowęglanowe) czy wybiórcze kanały jonowe – regulujące przepływ określonych jonów zgodnie z gradientem elektrochemicznym za pomocą otwierających się i zamykających bramek. Z kolei niektóre związki drobnocząsteczkowe (np. glukoza czy aminokwasy) podążają wtórnie za Na+, zgodnie z własnym gradientem stężeń. W błonach komórkowych znajdują się też receptory dla różnorakich mediatorów, których oddziaływanie zmienia przepływ określonych jonów.
Generalnie – błony komórkowe to bardzo wyspecjalizowany element naszego organizmu.
* *
Podsumowując:
Niniejsze opracowanie jest tylko ogólnym spojrzeniem na same podstawy gospodarki wodno-mineralnej. Chodzi o to, abyśmy uzmysłowili sobie, że równolegle z zasadą izotnii, drugim prawem homeostazy – prawem, którego nie może ona ignorować – jest zasada elektroobojętności: w każdej przestrzeni organizmu suma ładunków ujemnych dąży do zrównania się z sumą ładunków dodatnich (i vice versa).